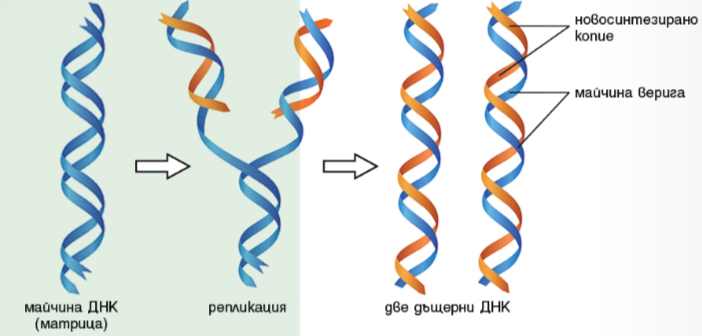
Способността за възпроизвеждане (авторепродукция) е основно свойство на живата материя. На молекулно ниво авторепродукцията се проявява чрез процеса репликация – процес на удвояване на молекулите на ДНК (удвояване на генетичната информация на клетката).
Репликация
Обща характеристика
Репликацията се извършва преди деленето на клетката. Всяка верига от майчината ДНК е матрица за синтеза на нова верига ДНК. Получените две дъщерни ДНК молекули са еднакви помежду си – всяка от тях има верига от майчината ДНК и новосинтезирана комплементарна верига. Така чрез репликацията се осигурява удвояване на генетичната информация, която по време на деленето се разделя поравно между двете дъщерни клетки.
Както всеки анаболитен процес, репликацията изисква енергия и специфични ензими. Енергията се доставя от предварително свързани с АТФ дезоксирибонуклеотиди. Активираните дезоксирибонуклеотиди са свързани не с един, а с три фосфорни остатъка, и притежават богати на енергия макроергични връзки. За протичането на репликацията е необходим и комплекс от ензими и белтъци, наречен белтъчен комплексна репликацията.
Механизъм на процеса
Репликацията започва с разделянето на двете комплементарни вериги на ДНК. Ензимът топоизомераза разсуква двойната спирала на ДНК и премахва спирализацията чрез разкъсване и последващо зашиване на една от веригите. Водородните връзки между двете вериги се разкъсват от ензими хеликаза. Предотвратяването на последваща спирализация на веригите се осигурява от т. нар. дестабилизиращи белтъци. Двете вериги се отделят една от друга, като се образува Y-образна структура, наречена репликационна вилка. Получената репликационна вилка напредва по дължината на ДНК молекулата.
Поради еднопосочността на действие на ДНК полимеразата (от 5’ към 3’ края) само върху една от веригите се осъществява непрекъснат синтез на нова верига. Активираните дезоксирибонуклеотиди се подреждат комплементарно върху ДНК веригата, завършваща с 3’ край. Ензимът ДНК полимераза ги свързва чрез 3’- 5’- фосфодиестерни връзки в полинуклеотидна верига. Веригата се синтезира бързо (от 5’ към 3’) и се нарича водеща верига.
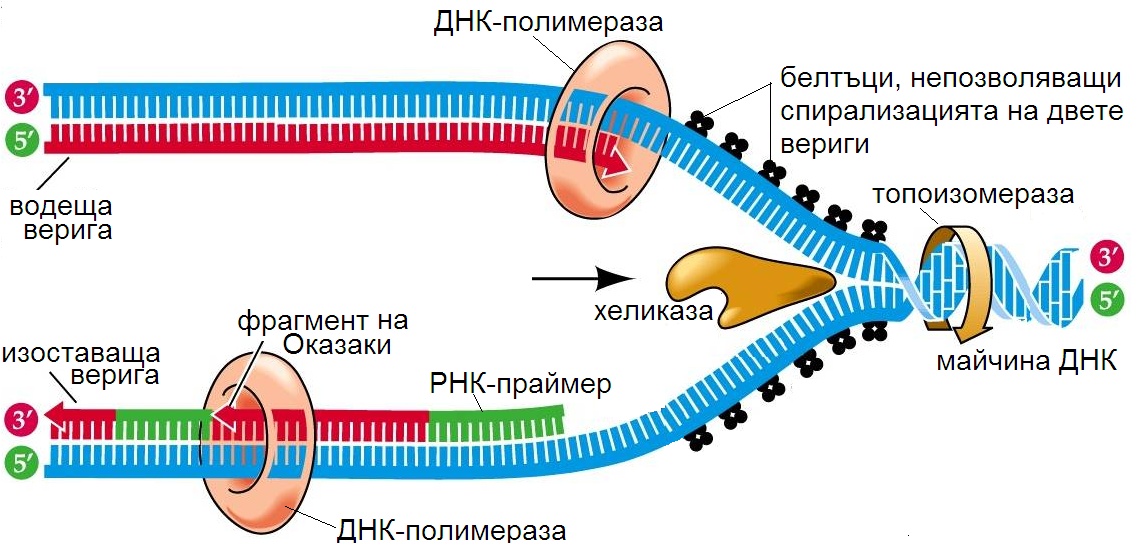
Синтезът на другата верига се извършва също от ензима ДНК полимераза, но на части (т. нар. фрагменти на Оказаки) в посока 5’- 3’. Тъй като ДНК полимеразата не може да започне синтеза от произволно място върху матрицата, първо се синтезира РНК зародиш от ензима РНК праймаза , към който ДНК полимеразата добавя нуклеотиди. Ензимът ДНК лигаза свързва фрагментите чрез 3’- 5’-фосфодиестерни връзки. Тази верига се синтезира по-бавно, поради което е наречена изоставаща.
Репликацията при еукариотните клетки е сравнително бавен процес – свързват се едва 10 до 40 нуклеотида в секунда. Причината за ниската скорост са хистоновите белтъци, около които е навита ДНК в хроматина. На практика репликацията не започва само от единия край на ДНК, а едновременно възникват голям брой начални точки на репликация, като се образуват многобройни реплициращи се участъци (репликони). Поради липса на структурни белтъци репликацията при прокариотите протича сравнително бързо – около 800 – 850 нуклеотида в секунда. Процесът се извършва, без да се нарушава пръстеновидната форма на ДНК молекулата.
Точността, с която се извършва процесът репликация, е много висока – около 1 грешка на 109 нуклеотидни двойки. Независимо от това за съхраняване на стабилността на ДНК молекулата еволюционно са възникнали механизми за коригиране на грешките – т. нар. репарация (поправка) на ДНК. Репарацията се извършва или веднага при допускането на грешката (в хода на репликацията), или след приключването на процеса (от ензима ДНК репараза).
За разлика от друга анаболитни процеси, които имат значение за клетката само докато тя съществува, и крайните им продукти не се предават в потомството, ДНК се синтезира само когато на клетката предстои делене, за да може клетъчната програма да се предаде непроменена в дъщерните клетки. Грешките при този синтетичен процес могат да доведат до смъртта на клетката или до мутации в нея.